
Angomonas deanei -Angomonas deanei
| Angomonas deanei | |
|---|---|
|
|
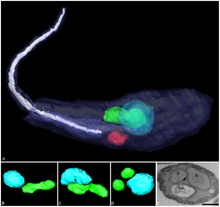
| Dreidimensionale Rekonstruktion von Angomonas deanei mit einem bakteriellen Endosymbionten (grün) in der Nähe seines Zellkerns (blau). | |
| Wissenschaftliche Klassifikation | |
| Domain: | |
| (ohne Rang): | |
| Stamm: | |
| Klasse: | |
| Befehl: | |
| Gattung: |
Angomonas
|
| Spezies: |
deanei
|
| Binomialer Name | |
|
Angomonas deanei (Carvalho 1973)
Teixeira & Camargo 2011 |
|
| Synonyme | |
|
Crithidia deanei Carvalho 1973 |
|
Angomonas deanei ist ein begeißeltes Trypanosomatid- Protozoon . Als obligater Parasit infiziert er den Magen-Darm-Trakt von Insekten und ist wiederum ein Wirt für symbiotische Bakterien. Der bakterielle Endosymbiont unterhält eine permanente wechselseitige Beziehung mit dem Protozoon, so dass er sich nicht mehr allein reproduzieren und überleben kann. Die Symbiose, die später in unterschiedlichem Maße auch bei anderen Protisten wie Strigomonas culicis , Novymonas esmeraldas , Diplonema japonicum und Diplonema aggregatum entdeckt wurde, gilt als gute Modelle für das Verständnis der Evolution von Eukaryoten aus Prokaryoten und der Entstehung von Zellorganellen (z Symbiogenese ).
Die Art wurde erstmals 1973 von der brasilianischen Parasitologin Aurora LM Carvalho als Crithidia deanei beschrieben . Eine phylogenetische Analyse im Jahr 2011 ergab, dass sie zur Gattung Angomonas gehört und damit zu Angomonas deanei wird . Das symbiotische Bakterium ist ein Mitglied des β-Proteobakteriums , das vom gemeinsamen Vorfahren mit der Gattung Bordetella oder wahrscheinlicher Taylorella abstammt . Die beiden Organismen sind so stark voneinander abhängig, dass sich das Bakterium nicht mehr vermehren kann und die Einzeller Insekten nicht mehr infizieren können, wenn sie isoliert sind.
Entdeckung
Angomonas deanei wurde ursprünglich als Crithidia deanei beschrieben . 1973 entdeckte die brasilianische Doktorandin Aurora Luiza de Moura Carvalho von der Universidade Federal de Goiás die Art aus seiner Studie über Darmparasiten der Killerwanzen in Goiás. Im nächsten Jahr berichtete er, dass der von ihm entdeckte Käfer Zelus leucogrammus nicht auf natürliche Weise mit dem Protozoen infiziert war, sondern von anderen Insekten erworben wurde. Gleichzeitig berichtete ein Forschungsteam der Universidade de Brasilia anhand der Transmissionselektronenmikroskopie über die biochemischen Eigenschaften und strukturellen Details. Sie entdeckten, dass es einen Endosymbionten beherbergt und beschrieben ihn als "wahrscheinlich bakteriell", der die "wichtigen Trypanosomatiden-Nährstoffe" lieferte. Die bakterielle Natur des Endosymbionten wurde 1977 bestätigt, als gezeigt wurde, dass er durch Behandlung mit dem Antibiotikum Chloramphenicol abgetötet werden kann und dass er dem Wirt bei der Synthese der Aminosäure Arginin aus Ornithin hilft.
Je mehr strukturelle und molekulare Details untersucht wurden, desto deutlicher wurde die Unterscheidung von A. deanei von anderen Crithidia- Arten. 1991 schlugen Maria Auxiliadora de Sousa und Suzana Corte-Real vom Instituto Oswaldo Cruz eine neue Gattung Angomonas für die Art vor. Eine phylogenetische Studie von Marta MG Teixeira und Erney P. Camargo von der Universität von São Paulo mit ihren Mitarbeitern im Jahr 2011 validierte den neuen Artnamen A. deanei zusammen mit einer Beschreibung einer neuen verwandten Art A. ambiguus , die auch die gleichen bakteriellen Endosymbionten enthält .
Struktur

Der Körper von Angomonas deanei hat eine elliptische Form mit einem markanten schwanzartigen Flagellum an seinem hinteren Ende zur Fortbewegung. Der bakterielle Endosymbiont befindet sich in seinem Körper und ist von zwei für Gram-negative Bakterien typischen Zellmembranen umgeben , aber seine Zellmembran weist ungewöhnliche Merkmale auf, wie das Vorhandensein von Phosphatidylcholin , einem wichtigen Membranlipid (atypisch für Bakterienmembranen) und der hochgradig reduzierte Peptidoglycan- Schicht, die eine reduzierte oder fehlende starre Zellwand zeigt . Die Zellmembran des Protozoen-Wirts enthält ein 18-Domänen- β-Barrel- Porin , das ein charakteristisches Protein von Gram-negativen Bakterien und ungewöhnlich für Eukaryoten ist. Darüber hinaus enthält es Cardiolipin und Phosphatidylcholin als die wichtigsten Phospholipide , während Sterine fehlen. Cardiolipin ist ein typisches Lipid von Bakterienmembranen; Phosphatidylcholin hingegen kommt hauptsächlich in symbiotischen Prokaryonten eukaryontischer Zellen vor. Zur symbiotischen Anpassung hat der Protozoen-Wirt Veränderungen erfahren, wie zum Beispiel reduzierte paraflagelläre Stäbchen, die für die volle Beweglichkeit der bakteriellen Geißeln erforderlich sind. Das paraflagellare Stäbchen-Gen PFR1 ist jedoch voll funktionsfähig. Es fehlen auch Introns und die Transkription langer polycistronischer mRNAs, die von anderen Eukaryoten für komplexe Genaktivitäten benötigt werden. Sein gesamtes Genom ist auf 29 Chromosomen verteilt und enthält 10.365 proteinkodierende Gene, 59 Transfer-RNAs, 26 ribosomale RNAs und 62 nicht-kodierende RNAs.
Während das Protozoon seine separaten Mitochondrien hat, die ein Elektronentransportsystem für die Produktion von zellulärer Energie bereitstellen, werden die ATP- Moleküle durch seine Glykosomen produziert . Es ist bekannt, dass das Bakterium den Wirt mit essentiellen Nährstoffen versorgt. Es synthetisiert Aminosäuren, Vitamine, stickstoffhaltige Basen und Häm für die Einzeller. Häm ist für das Wachstum und die Entwicklung des Protozoen notwendig. Das Bakterium stellt auch die Enzyme für den Harnstoffzyklus bereit, die im Wirt fehlen. Im Gegenzug bietet das Einzeller seine Enzyme für die kompletten Stoffwechselwege zur Biosynthese von Aminosäuren , Lipiden und Nukleotiden an , die im Bakterium fehlen. Das Bakterium hat im Vergleich zu seinen verwandten Bakterienarten ein stark reduziertes Genom, dem viele für sein Überleben notwendige Gene fehlen. Phosphatidylinositol , ein Membranlipid, das für die Zell-Zell-Interaktion in den Bakterien benötigt wird, wird ebenfalls von den Protozoen synthetisiert. Das Bakterium hängt auch für seine energetischen Funktionen vom Wirt für ATP-Moleküle ab. Somit teilen und tauschen die beiden Organismen ihre Stoffwechselsysteme innig miteinander aus.
Wenn das Bakterium mit Antibiotika abgetötet wird , kann das Protozoon aufgrund des veränderten Glycosylphosphatidylinositols (gp63) im Protozoengeißel keine Insekten mehr infizieren . Ein bakterienloses Protozoon weist reduzierte Genaktivitäten auf; insbesondere diejenigen, die am Oxidations-Reduktionsprozess beteiligt sind, werden der ATP-Hydrolyse-gekoppelte Protonentransport und die Glykolyse gestoppt. Die strukturellen Komponenten werden ebenfalls verändert, einschließlich der Zelloberfläche, der Kohlenhydratzusammensetzung, des paraflagellaren Stäbchens und des Kinetoplasten.
Schmarotzertum
Angomonas deanei wurde ursprünglich im Verdauungstrakt des Käfers Zelus leucogrammus entdeckt . Es wurde jedoch festgestellt, dass die Käfer nicht stark infiziert sind und wahrscheinlich von anderen Insekten übertragen wurden. Es ist nun bekannt, dass es verschiedene Moskitos und Fliegen infiziert und in der Lage ist, unter experimentellen Bedingungen Fibroblastenzellen von Säugetieren zu infizieren . Die Übertragung von einem Insekt zum anderen erfolgt nur zwischen Erwachsenen ( horizontale Übertragung ) und die Einzeller können sich nicht im Hinterdarm von Insektenlarven festsetzen. Das Flagellum wird als Klebeorgan verwendet, das in der Nähe der Rektumdrüsen und manchmal direkt auf der Oberfläche der Rektumdrüsen befestigt wird.
Reproduktion

Die zelluläre Reproduktion zeigt eine starke synergistische Anpassung zwischen dem Bakterium und dem Protozoon. Das Bakterium teilt sich zuerst, gefolgt von den Protozoen-Organellen und zuletzt dem Zellkern . Als Ergebnis enthalten die Tochterprotozoen genau die gleichen Kopien der Organellen und des bakteriellen Endosymbionten. Die gesamte Reproduktion dauert in einem idealen Nährmedium ca. 6 Stunden; Somit kann ein einzelnes Protozoon an einem Tag 256 Tochterzellen produzieren, obwohl es sich unter seinem natürlichen Lebensraum geringfügig unterscheiden kann.
Der Endosymbiont und die Evolution
Symbiotische Bakterien in den Trypanosomatiden-Protozoen stammen von einem β-Proteobakterium der Gattung Bordetella ab . Mit A. deanei , haben die Bakterien -co entwickelte sich in einer mutualistic Beziehung , die durch intensiven Stoffwechselaustausch gekennzeichnet ist . Der Endosymbiont enthält Enzyme und metabolische Vorläufer, die wesentliche Biosynthesewege des Wirts-Protozoen vervollständigen, wie die im Harnstoffzyklus und die Produktion von Hämin und Polyamin .
Das symbiotische Bakterium gehört zur Familie der β-Proteobakterien Alcaligenaceae . Basierend auf den 16S rRNA-Gensequenzen ist bekannt, dass es von einem gemeinsamen Vorfahren Kinetoplastid mit Blastocrithidia- Spezies stammt. Es wird angenommen, dass die beiden Gruppen in zwei verschiedene Wirtsprotozoen eindringen, um sich zu verschiedenen Arten zu entwickeln. Daher wurde dem Bakterium der wissenschaftliche Name ( Candidatus ) Kinetoplastibacterium crithidii gegeben. Obwohl ursprünglich vermutet wurde, dass das Bakterium aus einem gemeinsamen Vorfahren mit Mitgliedern von Bordetella hervorgegangen ist, ergab eine detaillierte phylogenomische Analyse , dass es enger mit Mitgliedern der Gattung Taylorella verwandt ist .